Riž. 4.21. Položaj električne osi srca je vodoraven (kot a * +15*). Obstaja tudi rotacija srca okoli vzdolžne osi v nasprotni smeri urinega kazalca (zapleteno QRS v vodih tipa V 5 in V 6 QR, prehodno območje (PZ) v vodi V 2.
Riž. 4.22. Obstaja vrtenje električne osi srca v desno (kot a * + 120 °), pa tudi vrtenje srca okoli vzdolžne osi v smeri urinega kazalca do PZ v vodi V 6 (kompleksno QRS v vodih V 5 (V 6 tipa AS).
Določanje vrtenja srca okoli prečne osi (na vrhu naprej ali nazaj)
Manj pogosto EKG beleži rotacije srca okoli njegove prečne osi, ki se pojavljajo v anteroposteriorni (sagitalni) ravnini (slika 4.23). Rotacije srca okoli prečne osi so običajno povezane z odstopanjem vrha srca naprej ali nazaj vzdolž
glede na njegov običajni položaj, kar vodi v kršitev običajne prostorske lege treh momentnih vektorjev ventrikularne depolarizacije v sagitalni in čelni ravnini. Vrtenje srca naprej ali nazaj okrog prečne osi je najbolje zabeležiti v treh standardnih vodih okončin. Oglejte si sl. 4.23. Prikazuje znani šestosni Baileyjev koordinatni sistem, vrnjen pod določenim kotom na opazovalca, pa tudi prostorsko lokacijo treh vektorjev trenutka (0,02 s, 0,04 s in 0,06 s).
V večini primerov je pri normalnem položaju srca (slika 4.23, a) vektor začetnega momenta (0,02 s) nekoliko navzgor in desno, vektor končnega trenutka (0,06 s) pa navzgor in levo ali desno. Oba vektorja sta prostorsko nameščena pod določenim kotom na čelno ravnino, pri čemer je vektor 0,02 s usmerjen naprej, vektor 0,06 s pa nazaj. Oba vektorja sta projicirana na negativne dele osi standardnih vodnikov, zaradi česar je v teh vodih mogoče zabeležiti relativno majhne amplitude valov Q in Q. S. Ne smemo pozabiti, da zobje Vprašanje in S jih je mogoče zabeležiti le v enem ali dveh od treh standardnih odvodov: v I in II ali v II in III.
nasvet naprej(Slika 4.23, b) se vektor začetnega momenta (0,02 s) premakne še bolj navzgor in nekoliko v desno, zato je zob Vprašanje se začne registrirati v vseh treh standardnih vodih in postane bolj izrazit.
Vektor končnega momenta (0,06 s) se odkloni navzdol in nazaj, zaradi česar se zdaj nahaja skoraj pravokotno na čelno ravnino. Zato se njegova projekcija na os vseh standardnih vodov približa ničli, kar vodi do izginotja vala 5 v teh vodih.
Ko se srce obrne okoli prečne osi nasvet nazaj(Slika 4.23, c) se vektor začetnega momenta (0,02 s) premakne naprej in navzdol, tako da se izkaže, da je njegova orientacija v prostoru skoraj pravokotna na čelno ravnino. Zato se projekcija vektorja 0,02 s na os standardnih vodov približa ničli, sami zobje Vprašanje niso registrirani.
Končni vektor navora (0,06 s) se premakne še bolj navzgor in se začne projicirati na negativne osi vseh treh standardnih okončin, kar vodi v pojav dovolj globokih zob S v S u in S m
Tako je za določitev vrtenja srca okoli prečne osi treba oceniti konfiguracijo kompleksa QRS v standardnih vodih okončin.
Analiza atrijskega vala R
Po določitvi rotacij srca okoli anteroposteriorne, vzdolžne in prečne osi nadaljujte z analizo atrijskega zoba R. Analiza zobnikov R vključuje: 1) merjenje amplitude vala R, 2) merjenje trajanja vala PŽP 3) določitev polarnosti zoba R, 4) določanje oblike zobnika R.
Amplituda valov R se meri od izolinije do vrha vala in traja od začetka do konca vala, kot je prikazano

na sl. 4.24. Normalna amplituda valov R ne presega 2,5 mm in traja 0,1 s. Polarnost vala R v vodih I, II in III je najpomembnejši elektrokardiografski znak, ki označuje smer premikanja vzbujalnega vala vzdolž atrija in posledično lokacijo vira vzbujanja (srčni spodbujevalnik). Kot se spomnite, so pri normalnem gibanju vzbujalnega vala vzdolž atrijev od zgoraj navzdol in levo zobje /> pozitivni, pri vzbujanju pa od spodaj navzgor negativno. srčni spodbujevalnik se nahaja v spodnjem atriju ali v zgornjem delu AV vozla. Z vzburjenostjo, ki izvira iz srednjega dela desnega atrija, je val depolarizacije usmerjen navzgor in navzdol. Srednji vektor R usmerjen v levo oziroma zob R ( narašča, postaja vse bolj izrazit P lv zob P || (postane negativen plitv.
Določanje oblike zoba je zelo praktičnega pomena. R. Razcepljen z dvema vrhovoma, razširjenim zobom R v levih vodih (I, aVL, V 5, V 6) je značilno za bolnike z mitralnimi srčnimi napakami in hipertrofijo levega atrija ter koničaste zobe z visoko amplitudo R v odvodih I, III, aVF opazimo pri hipertrofiji desnega atrija pri bolnikih s srčnim pulmom (za več podrobnosti glejte 7. poglavje).
Analiza ventrikularnega kompleksa QRST
Za poddružino pravega šaša (Caricoideae) so značilni enospolni cvetovi, ki nimajo cvetočega, razen sibirske kobrezije (Kobresia sibirica), v kateri je sestavljena iz treh rjavih lusk (slika 167). Kosički v predstavnikih te družine sedijo v pazduhah luskastih pokrivnih listov in so obdani z luskami, ki se nahajajo na ventralni (trebušni) strani klasca in so homologne s predlistom vegetativnega poganjka. V rodovih Schoenoxiphium in Kobresia ima ta lestvica nenarasle ali do polovice (redko do vrha) spojene robove, v rodovih Uncinia in šaš (Carex) pa je močno metamorfozirana, zaprta, vrečasto podobna. Šenoksifi in kobrezija imajo običajno dvospolne klasčke s podolgovato (v prvem rodu) in močno skrajšano, komaj razvito (v drugem rodu) os. V unciniji in šašu so klasčki zmanjšani na en sam ženski cvet, zaprt v vrečkasto lusko, imenovano vrečka. Pri vrsti uncinia ženski cvet sedi na dnu še ohranjene osi konice; slednji se pri veliki večini šašev popolnoma zmanjša. Prašniki 3. Steber z 2 - 3 vejami stigme. Poddružino sestavlja eno pleme šaša (Cariceae), vključno s 4 rodovi. Najbolj primitiven med njimi je schenoxyphyum, ki šteje približno 15 vrst, od katerih 2 rasteta na Madagaskarju, preostali pa v Južni Afriki. Rastejo v gorah, na vlažnih in močvirnih krajih. To so trajnice z bazalnimi, ponavadi ploskimi listi in konicami ali paličastimi socvetji. Kosičke obkrožajo zaprte, v različni meri, včasih vrečaste luske. Znotraj nje je močno sploščena os konica, na dnu katere sedi ženska roža, na vrhu pa je več moških (slika 167). Pri nekaterih vrstah os ne nosi moških cvetov; slednji v istem socvetju tvorijo samčke.
Naslednji rod poddružine šašev, Ouncinia, sestavlja 40-50 vrst, razširjenih predvsem na južni polobli (razen v Afriki). Večina vrst (32) izvira iz Nove Zelandije. Severno od ekvatorja uncinia najdemo v jugovzhodni Aziji (otok Kalimantan, Filipini), na Havajih, v Srednji in Južni Ameriki ter v Zahodni Indiji. Vrste tega roda rastejo od morske gladine do alpskega pasu, na močvirnih travnikih, močvirjih, v vlažnih gozdovih in grmovju, kjer pogosto tvorijo goščave. Vrsta Ouncinia je trajnica, običajno gosto travnata trava z bazalnimi linearnimi listi. Socvetje je apikalni klas, ki v zgornjem delu nosi moške cvetove, v spodnjem delu pa vrečke, ki obdajajo ženske cvetove, iz katerih na vrhu štrli os kljukaste oblike. Vrh osi v obliki kljuke je toga, ovita pokrivna luska, v sinusu katere je, kot kažejo anatomske študije, nerazvita ledvica. Prašniki 3. Steber s 3 stigmatiznimi vejami. Plod je trikoten, zaprt v vrečko. S kljukasto upognjeno osjo se vrečke oprimejo volne ali perja živali in se tako razširijo.
Rod Kobresia obsega približno 40 vrst, razširjenih predvsem v visokogorju zmernih in deloma subtropskih in tropskih pasov severne poloble, predvsem pa v Aziji; nekaj vrst prodira na Arktiko. Kobrezija je značilna gorska rastlina z malo padavinami. Rastejo ob bregovih rek in jezer, na vlažnih in močvirnih, pogosto slanih travnikih, travniško-stepskih in skalnatih pobočjih. Vrste tega rodu pogosto tvorijo kobrezijske travnike (tako imenovane kobreznike ali puščave). Kobresia - trajna gosta travnata trava z bazalnimi listi, ki imajo ravne ali ščetinaste plošče. Socvetje je v obliki konic ali, manj pogosto, v metlici, sestavljeno iz majhnih 2-7-cvetnih dvospolnih klasic, od katerih so zgornji včasih moški. Luske, ki obkrožajo konice, imajo običajno neraste ali, redko, do polovice ali skoraj do vrha priraslih robov. Bodice (njihova os je močno skrajšana, skoraj ne izražena) so sestavljene iz enega ženskega cvetja in 1 - 6 moških cvetov, ki se nahajajo zunaj njega. Slednji so sestavljeni iz 3 prašnikov, ki sedijo v osi pokrivne luske. Steber s 3, redko 2 stigma vejama. Plod je običajno trikoten. Kobresia se razmnožuje s semeni, le eno izmed njih - močna kobresia (K. robusta) - pa tudi vegetativno, s pomočjo plazeče korenike. Plodove širijo rastlinojedi sesalci. Dlakava kobrezija (K. capilliformis), ki zavzema velika območja v Tien Shanu in Pamir -Alaju, pa tudi številne druge vrste - dragocene pašnike.

Rod šaša (Carex) ima približno 1500 (po drugih virih do 2000 - 2500) vrst in spada v število relativno majhnih največjih rodov kritosjemenk. Vrste šaša so razširjene po vsem svetu, predvsem pa v zmernih in hladnih regijah, kjer imajo pomembno vlogo pri sestavi rastlinskega pokrova ne le v mokrih in močvirnih, ampak tudi v suhih habitatih. Po udeležbi v rastlinskem pokrovu Sovjetske zveze, kjer najdemo približno 400 vrst šašev, so na drugem mestu po travah in slonokosih. Mnoge vrste tega rodu so edifikatorji različnih rastlinskih skupnosti, vključno s stepe in celo polpuščavo in puščavo. Šaši rastejo skoraj povsod, od morske gladine do visokogorja. Večina vrst naseljuje močvirja, močvirne in vlažne travnike, obale vodnih teles (na primer steklenica - C. rostrata, mehurček - C. vesicaria, lisica - C. vulpina, slika 174, dvodomna šaša - C . dioica, riž. 174). Druge šaše najdemo v različnih vrstah gozdov, tudi v tropskih deževnih gozdovih. Nekatere vrste rastejo na suhih travnikih, stepah, suhih pobočjih (nizek šaš - C. hnmilis, zgodnji šaš - C. praesokh, počep - C. supina itd.). Pachystylis šaš (C. pachystylis) je značilna rastlina glinastih polpuščav, medtem ko je otekla šaša (C. physodes) značilna rastlina peščenih puščav. Slednja vrsta ima nenavadno izviren videz zaradi močno mehurjastih otečenih, rdečkasto zarjavelih, zelo velikih (do 2 cm dolgih) vreč (slika 174). Kar nekaj vrst živi ob morskih obalah, peščenih sipinah, obmorskih travnikih, prodnikih, skalah. Večina vrst v zmernem pasu raste v ravninah, nekateri šaši pa so tipični prebivalci subalpskih in alpskih travnikov. V tropih, zlasti v malezijski floristični regiji, kjer raste 66 vrst šašev, slednje živijo predvsem v gorah in le približno 10 vrst raste od morske gladine do 500 m nad morjem.
Šaš je večletna enoedna, zelo redko dvodomna trava s plazečo ali skrajšano koreniko, ki tvori trate ali včasih izbokline. Listi so običajno vsi bazalni, veliko manj pogosto so tudi stebelni listi, ki segajo od vozlišč, ki se nahajajo skoraj vzdolž celotnega stebla; listna plošča je običajno linearna, redko suličasta (na primer pri rjasto peščeni šaši - C. siderosticta, slika 175), včasih pa zožena v peclje. Cvetovi so enospolni, z klasčki, ki tvorijo klasaste, metličaste, grozdaste ali glavaste socvetje; včasih je socvetje enojno. Kosički so v celoti sestavljeni iz moških ali ženskih cvetov ali so mešani, dveh vrst: androgini - z moškimi cvetovi v zgornjem delu klasca in ženski - v spodnjem, ginekandrični - z ženskimi cvetovi na vrhu, z moškimi cvetovi na dnu . Ženski cvetovi so zaprti v vrečko, ki raste s plodovi, na vrhu zoženi v izliv, iz katerega so izpostavljeni del stebra in vejice stigme. Zgoraj je bilo že povedano, da so ženski cvetovi šaša, zaprti v vrečah, zmanjšani, enocvetni klasci. Zato je zbirka vrečk, ki sedijo na eni osi, zapleteno uho. Zbirke istih moških cvetov so preprosta ušesa. Kljub temu je zaradi udobja običajno zapletene in preproste šopke šaša preprosto imenovati šopke. Znaki vrečk so zelo raznoliki in so zelo pomembni za taksonomijo šašev. Številne vrste tega rodu se razlikujejo predvsem po vrečah. Vrečke so organ nedvomnega biološkega pomena. Morda je zaradi videza vreče rod šaša postal najštevilčnejši in najbolj razširjen od vseh rodov družine. Vrečke ščitijo jajčnik in plod v razvoju pred neugodnimi razmerami, pri mnogih vrstah pa so tudi prilagoditev za širjenje plodov. Steblo v šašu z 2 - 3 običajno dolgimi vejami. Plodovi so trikotni ali rahlo izbočeni.
Številni šaš cveti zgodaj spomladi. Vrste tega rodu so anemofilne rastline, nekatere pa očitno imajo tudi entomofilijo. Tako je znano, da dve najzgodnejši cvetoči vrsti - vresin šah (C. ericetorum) in klinček (C. caryophyllea) - redno obiskujeta čebele za nabiranje cvetnega prahu, ki očitno izvajata navzkrižno opraševanje. Podatkov o biologiji cvetenja šaša skoraj ni. Njihovi cvetovi veljajo za protogene. Toda, kot kažejo posebna opazovanja cvetenja dlakavega šaša (C. pilosa), morfološka protoginija morda ne sovpada s fiziološko (L. Antonova, 1976). Pri tej vrsti je videz popolnoma razvitih stigm pred odpiranjem prašnikov. Vendar pa stigme dosežejo fiziološko zrelost, to je sposobnost zaznavanja cvetnega prahu, šele takrat, ko se prašniki odprejo in se cvetni prah razprši. Tako dlakavi šaš v bistvu ni prototip, ampak homogamna rastlina. Samoopraševanje je verjetno izključeno zaradi delne samo-nezdružljivosti. Eksperimentalno je bilo ugotovljeno, da pri samoopraševanju pri tej vrsti nastane bistveno manj plodov kot pri navzkrižnem opraševanju. Šaši se razmnožujejo s semeni, pri vrstah s plazečimi koreniki pa prevladuje vegetativno razmnoževanje. Do širjenja sadja pride na različne načine. Plodovi obalnih šašev, zaprti v nabreklih ali gobastih vrečah na dnu, se širijo po vodi. Vreče otekle šaše so dobro prilagojene anemohoriji. Z lahkoto se valjajo po površini peska, tudi pri rahlem vetru. Pri mnogih vrstah se vrečke širijo epizooholično, predvsem z umazanijo na šapah ptic. Vrečke nekaterih šašev se lahko oprimejo tudi perja ptic. Pomembno vlogo ima endozojska porazdelitev plodov, ko jih pogoltnejo race. Ptice nosijo oranžno rdeče, nekoliko mesnate vrečke tropske vrste jagodičaste šaše (C. baccans, slika 175). Vreče nekaterih gozdnih šašev, na primer prstne šaše (C. digitata) in perutninske šaše (C. ornithopoda), ki imajo podolgovate mesnate podlage, nosijo mravlje.
Manj pogosto EKG beleži rotacije srca okoli njegove prečne osi, ki se pojavljajo v anteroposteriorni (sagitalni) ravnini (slika 57). Rotacije srca okoli prečne osi so običajno povezane z odstopanjem vrha srca naprej ali nazaj glede na njegov običajni položaj, kar vodi v kršitev običajne prostorske lege treh momentnih vektorjev ventrikularne depolarizacije v sagitalne in čelne ravnine.
Rotacije srca okoli prečne osi z vrhom naprej ali nazaj je najbolje zabeležiti v treh standardnih vodih od okončin. Oglejte si sl. 57. Prikazuje znani šestosni Baileyjev koordinatni sistem, zasukan pod določenim kotom glede na opazovalca, ter prostorno lokacijo treh vektorjev trenutka (0,02 s, 0,04 in 0,06 s).
IN
Riž. 57. EKG oblika v treh standardnih odvodih. a - z normalnim položajem srca, b - ko je srce obrnjeno okoli prečne osi z vrhom naprej; c - ko je srce obrnjeno okoli prečne osi z vrhom nazaj.
Oba vektorja sta prostorsko nameščena pod določenim kotom na čelno ravnino, vektor 0,02 s je usmerjen naprej, vektor 0,06 s pa nazaj. Oba vektorja sta projicirana na negativne dele osi standardnih vodnikov, zaradi česar je v teh odvodih mogoče zabeležiti relativno majhne amplitude valov Q in S. Ne smemo pozabiti, da je valove Q in S mogoče zabeležiti le v enega ali dva od treh standardnih vodov: v I in II ali v II in III.
Ko se srce obrne okoli prečne osi z vrhom naprej (slika 57, b), se vektor začetnega momenta (0,02 s) premakne še bolj navzgor in nekoliko v desno, zato se val Q začne zapisovati v vseh treh standard vodi in postane bolj izrazit.
Vektor končnega trenutka (0,06 s) se odkloni navzdol in nazaj, zaradi česar se zdaj nahaja skoraj pravokotno na čelno ravnino. Zato se njegova projekcija na os vseh standardnih vodov približa ničli, kar vodi do izginotja vala S v teh vodih.
Spomnite se! Ko se srce vrti okoli prečne osi z vrhom naprej, ima ventrikularni kompleks QRS v standardnih vodih obliko qR I, qR II in qR III
Ko se srce obrne okoli prečne osi z vrhom nazaj (slika 57, c), se vektor začetnega momenta (0,02 s) premakne naprej in navzdol, tako da je njegova orientacija v prostoru skoraj pravokotna na čelno ravnino. Zato se projekcija vektorja 0,02 s na os standardnih vodov približa ničli, sami valovi Q pa se ne zabeležijo.
Končni vektor navora (0,06 s) se premakne še bolj navzgor in se začne projicirati na negativne osi vseh treh standardnih okončin, kar vodi v pojav dovolj globokih zob S I, S II in S III.
Spomnite se! Ko se srce obrne okoli prečne osi z vrhom nazaj, ima ventrikularni kompleks v standardnih vodih obliko RS I, RS II, RS III.
Tako je za določitev zavojev srca okoli prečne osi treba oceniti konfiguracijo kompleksa QRS v standardnih vodih iz okončin.
Riž. 58. Merjenje amplitude in trajanja vala P EKG. A p je amplituda vala P, t p je trajanje vala P.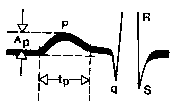
Socvetje je poganjki ali sistem poganjkov, ki nosijo cvetove. Na vozliščih osi socvetja se nahajajo isti listi kot na vegetativnem delu poganjka ali spremenjeni listi, ki so izgubili sposobnost fotosinteze - brakteje, in na vozliščih pedikela - brakteje.
Biološka prednost socvetij pred posameznimi cvetovi je povečati jamstvo opraševanja, zmanjšati verjetnost poškodb cvetov zaradi neugodnih okoljskih dejavnikov zaradi njihovega postopnega cvetenja. Večina rastlin ima socvetja.Obstajata dve vrsti socvetij: zapleteno ko se cvetovi nahajajo na vejah glavne osi, in preprosto ko se cvetje z ali brez pecljev nahaja neposredno na glavni osi.
Kompleksna socvetja, odvisno od načina rasti osi, delimo na: simpodial(določeno) - os se konča v cvetu, cvetenje cvetov poteka od vrha do stranskih vej ali centrifugalno, če se cvetovi nahajajo v isti ravnini; in monopodialni(nedoločeno) - os raste v nedogled, cvetovi cvetov gredo od podlage proti vrhu ali centripetalno, če se cvetovi nahajajo v isti ravnini.
Kompleksna simpodialna socvetja:
- monochasium - glavna os se konča s cvetjem; pod njim se oblikuje os drugega reda, tudi okronana s cvetjem itd. če se podcvetne osi premikajo v eno smer, se tvori zavoj, če pa izmenično v eno smer, potem v drugo smer, nastane girus; kodra, pri katerem se stranske veje skrajšajo, se imenuje glomerulus;
- dihazij - pod cvetom nastaneta dve nasprotni osi, od katerih se vsaka prav tako konča v cvetu in daje tudi dve podcvetni osi, ki ponavljata isto metodo razvejanja itd .;
- pleiochasium - od glavne osi, ki nosi en apikalni cvet, se oddaljujeta več kot dve podcvetni osi, ki tvorita vrtinec monohazije ali dihazije;
- thyrsus - zgornja socvetja se nahajajo na glavni osi, pogosteje imajo piramidalno obliko.
Kompleksna monopodialna socvetja:
- metlica - zelo razvejano socvetje, spodnje stranske razvejanosti se veje močneje od zgornjih;
- scutellum - metlica, v kateri se cvetovi nahajajo bolj ali manj v isti ravnini;
- kompleksna konica - glavna os tvori veje, na katerih se nahajajo cvetovi brez pecljev; te veje imenujemo klasci;
- kompleksen dežnik-razdalje med osmi drugega reda se skrajšajo in se odmikajo od vrha osi prvega reda; razdalje med peclji se skrajšajo in se nahajajo na vrhu osi drugega reda; pogosto listi na dnu osi drugega reda tvorijo skupno ovojnico, na dnu pecljev pa zasebno ovojnico.
Poleg naštetih obstajajo tudi kompleksna socvetja, imenovana agregat... Nastanejo z združevanjem različnih vrst socvetij. Na primer, rman ima agregatno socvetje - košare, zbrane v scutellum; močvirja in modrikavec imata v metlici zbrane klasice.
Preprosta socvetja s podolgovato osjo:
- čopič - glavna os nosi rože s peclji, običajno enake dolžine;
- konica - glavna os nosi rože brez pecljev;
- uhan - povešeno uho, to je uho z mehko osjo; po cvetenju socvetje običajno odpade;
- uho - uho z močno odebeljeno osjo, obdano z enim ali več listi, tako imenovano tančico ali krilom.
Preprosta socvetja s skrajšano osjo:
- dežnik - socvetje, v katerem je glavna os skrajšana, pedikeli, ki imajo skoraj enako dolžino, pa se odmikajo od vrha osi;
- glava je dežnik s cvetjem ali brez pecljev ali zelo kratek;
- košara - vrh glavne osi raste v obliki postelje, na kateri so tesno zaprti cvetovi; apikalni listi so natrpani in tvorijo ovoj.
Cimoidna socvetja- To je obsežna skupina socvetij. Med cimoidnimi socvetji obstajata dve glavni vrsti: cimoidi in tirsus... Cimoidi so praviloma poenostavljeni tirsus. Thyrsi so razvejana socvetja, stopnja razvejanja pa se zmanjšuje od podlage do vrha. Glavna os tirusa raste monopodialno, toda cimoidi so delna ali drugačna socvetja.
Thyrsus, katerega glavna os se konča v cvetu, imenujemo zaprti, sicer veljajo za odprte. Glede na stopnjo razvejanosti stranskih osi ločimo pleiotirje, pri katerih se cimoidi nahajajo na osi tretjega in višjega reda; ditiri, pri katerih so cimoidi na osi drugega reda; in monothyrsi, pri katerih se cimoidi nahajajo neposredno na glavni osi socvetja. Zunanja podobnost tirsusa z grozdjem, ušesom, uhanom, dežnikom ali glavo kaže na grozdaste, konaste, ušesne, dežnične, glavolične oblike tvoje itd. Thyrsi se z zmanjšanjem (zmanjšanjem) števila stranskih osi, skrajšanjem internodij itd. Zlahka spremenijo v druge vrste socvetij. Thyrsus so zelo pogosti pri rastlinah. Na primer, thrsus je socvetje divjega kostanja. Thyrsi različnih vrst so socvetja številnih labatov. Brezova socvetja so uhani tirsus.


